














 |
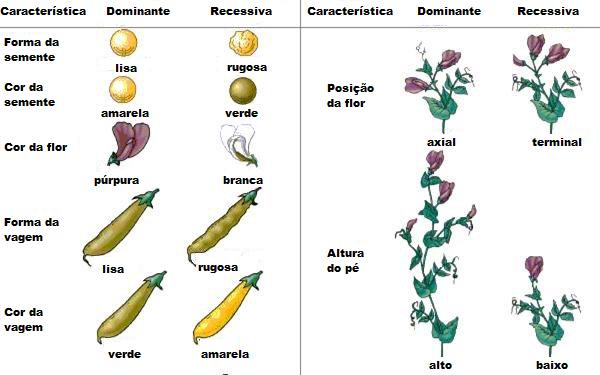
Mendel, o iniciador da genética
Após ter sido
ordenado monge, em 1847, Mendel ingressou na Universidade de Viena, onde
estudou matemática e ciências por dois anos. Ele queria ser professor
de ciências naturais, mas foi mal sucedido nos exames.
De volta a Brünn, onde passou o
resto da vida. Mendel continuou interessado em ciências. Fez estudos
meteorológicos, estudou a vida das abelhas e cultivou plantas, tendo
produzido novas variedades de maças e peras. Entre 1856 e 1865, realizou
uma série de experimentos com ervilhas, com o objetivo de entender como
as características hereditárias eram transmitidas de pais para filhos.
Em 8 de março de 1865, Mendel
apresentou um trabalho à Sociedade de História Natural de Brünn, no qual
enunciava as suas leis de hereditariedade, deduzidas das experiências
com as ervilhas. Publicado em 1866, com data de 1865, esse trabalho
permaneu praticamente desconhecido do mundo científico até o início do
século XX. Pelo que se sabe, poucos leram a publicação, e os que leram
não conseguiram compreender sua enorme importância para a Biologia. As
leis de Mendel foram redescobertas apenas em 1900, por três
pesquisadores que trabalhavam independentemente.
|
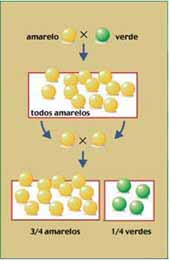 |
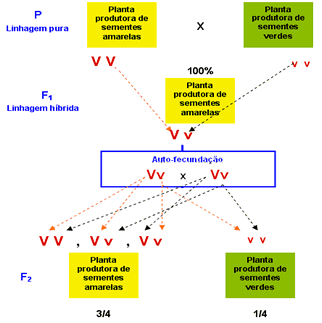
Após repetir o mesmo procedimento
diversas vezes, Mendel verificou que todas as sementes originadas
desses cruzamentos eram amarelas – a cor verde havia aparentemente
“desaparecido” nos descendentes híbridos (resultantes do cruzamento das
plantas), que Mendel chamou de F1 (primeira geração filial). Concluiu, então, que a cor amarela “dominava” a cor verde. Chamou o caráter cor amarela da semente de dominante e o verde de recessivo.
A seguir, Mendel fez germinar as sementes obtidas em F1
até surgirem as plantas e as flores. Deixou que se autofertilizassem e
aí houve a surpresa: a cor verde das sementes reapareceu na F2
(segunda geração filial), só eu em proporção menor que as de cor
amarela: surgiram 6.022 sementes amarelas para 2.001 verdes, o que
conduzia a proporção 3:1. Concluiu que na verdade, a cor verde das sementes não havia “desaparecido” nas sementes da geração F1.
O que ocorreu é que ela não tinha se manifestado, uma vez que, sendo
uma caráter recessivo, era apenas “dominado” (nas palavras de Mendel)
pela cor amarela. Mendel concluiu que a cor das sementes era determinada
por dois fatores, cada um determinando o surgimento de uma cor, amarela
ou verde.
|
VV
|
vv
|
Vv
|
Semente amarela pura
|
Semente verde pura
|
Semente amarela híbrida
|
 |
Em 1902, enquanto estudava a
formação dos gametas em gafanhotos, o pesquisador norte americano Walter
S. Sutton notou surpreendente semelhança entre o comportamento dos
cromossomos homólogos, que se separavam durante a meiose, e os fatores
imaginados por Mendel. Sutton lançou a hipótese de que os pares de
fatores hereditários estavam localizados em pares de cromossomos
homólogos, de tal maneira que a separação dos homólogos levava à
segregação dos fatores.
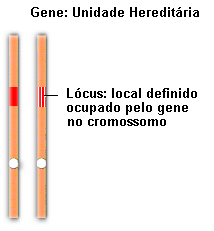
Hoje sabemos que os fatores a que Mendel se referiu são os genes (do grego genos,
originar, provir), e que realmente estão localizados nos cromossomos,
como Sutton havia proposto. As diferentes formas sob as quais um gene
pode se apresentar são denominadas alelos. A cor amarela e a cor verde
da semente de ervilha, por exemplo, são determinadas por dois alelos,
isto é, duas diferentes formas do gene para cor da semente.
|
B
Gametas paternos
b
|
Gametas maternos
B b
|
|
BB
Preto
|
Bb
Preto
|
|
Bb
Preto
|
bb
Branco
|
|
|
Os genes que condicionam a produção desses antígenos são apenas dois: L M e L N (a letra L é a inicial do descobridor, Landsteiner). Trata-se de uma caso de herança medeliana simples. O genótipo L ML M, condiciona a produção do antígeno M, e L NL N, a do antígeno N. Entre L M e L N há co-dominância, de modo que pessoas com genótipo L ML N produzem os dois tipos de antígenos.
|
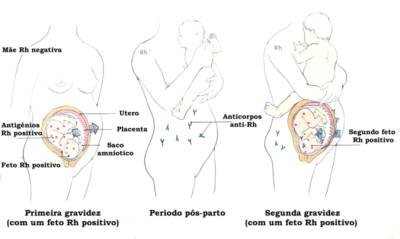
| Para simplificar, no entanto, considera-se o envolvimento de apenas um desses pares na produção do fator Rh, motivo pelo qual passa a ser considerado um caso de herança mendeliana simples. O gene R, dominante, determina a presença do fator Rh, enquanto o gene r, recessivo, condiciona a ausência do referido fator. |
|
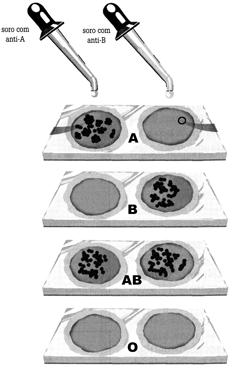
Determinação dos grupos sanguíneos
utilizando soros anti-A e anti-B. Amostra 1- sangue tipo A. Amostra 2 -
sangue tipo B. Amostra 3 - sangue tipo AB. Amostra 4 - sangue tipo O.
|
A descoberta dos grupos sanguíneos
Por volta de 1900, o médico austríaco Karl
Landsteiner (1868 – 1943) verificou que, quando amostras de sangue de
determinadas pessoas eram misturadas, as hemácias se juntavam, formando
aglomerados semelhantes a coágulos. Landsteiner concluiu que
determinadas pessoas têm sangues incompatíveis, e, de fato, as pesquisas
posteriores revelaram a existência de diversos tipos sanguíneos, nos
diferentes indivíduos da população.
Quando, em uma transfusão, uma pessoa recebe um
tipo de sangue incompatível com o seu, as hemácias transferidas vão se
aglutinando assim que penetram na circulação, formando aglomerados
compactos que podem obstruir os capilares, prejudicando a circulação do
sangue.
Aglutinogênios e aglutininas
No sistema ABO existem quatro tipos de sangues: A, B, AB e O.
Esses tipos são caracterizados pela presença ou não de certas
substâncias na membrana das hemácias, os aglutinogênios, e pela presença
ou ausência de outras substâncias, as aglutininas, no plasma sanguíneo.
Existem dois tipos de aglutinogênio, A e B, e
dois tipos de aglutinina, anti-A e anti-B. Pessoas do grupo A possuem
aglutinogênio A, nas hemácias e aglutinina anti-B no plasma; as do grupo
B têm aglutinogênio B nas hemácias e aglutinina anti-A no plasma;
pessoas do grupo AB têm aglutinogênios A e B nas hemácias e nenhuma
aglutinina no plasma; e pessoas do gripo O não tem aglutinogênios na
hemácias, mas possuem as duas aglutininas, anti-A e anti-B, no plasma.
|
ABO
|
Substâncias
|
%
|
Pode receber de
|
||||||||
Tipos
|
Aglutinogênio
|
Aglutinina
|
Frequência
|
A+
|
B+
|
A+
|
0+
|
A-
|
B-
|
AB-
|
O-
|
AB+
|
A e B
|
Não Contém
|
3%
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
A+
|
A
|
Anti-B
|
34%
|
X
|
X
|
X
|
X
|
||||
B+
|
B
|
Anti-A
|
9%
|
X
|
X
|
X
|
X
|
||||
O+
|
Não Contém
|
Anti-A e Anti-B
|
38%
|
X
|
X
|
||||||
AB-
|
Ae B
|
Não Contém
|
1%
|
X
|
X
|
X
|
X
|
||||
A-
|
A
|
Anti-B
|
6%
|
X
|
X
|
||||||
B-
|
B
|
Anti-A
|
2%
|
X
|
X
|
||||||
O-
|
Não Contém
|
Anti-A e Anti-B
|
7%
|
X
|
|||||||
Indivíduos do grupo A não podem
doar sangue para indivíduos do grupo B, porque as hemácias A, ao
entrarem na corrente sanguínea do receptor B, são imediatamente
aglutinadas pelo anti-A nele presente. A recíproca é verdadeira:
indivíduos do grupo B não podem doar sangue para indivíduos do grupo A.
Tampouco indivíduos A, B ou AB podem doar sangue para indivíduos O, uma
vez que estes têm aglutininas anti-A e anti-B, que aglutinam as hemácias
portadoras de aglutinogênios A e B ou de ambos.
Assim, o aspecto realmente importante da
transfusão é o tipo de aglutinogênio da hemácia do doador e o tipo de
aglutinina do plasma do receptor. Indivíduos do tipo O podem doar sangue
para qualquer pessoa, porque não possuem aglutinogênios A e B em suas
hemácias. Indivíduos, AB, por outro lado, podem receber qualquer tipo de
sangue, porque não possuem aglutininas no plasma. Por isso, indivíduos
do grupo O são chamadas de doadores universais, enquanto os do tipo AB
são receptores universais.
|
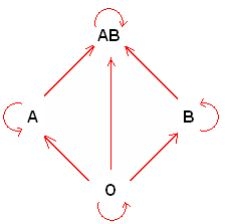 |
Fenótipos
|
Genótipos
|
A
|
I AI A ou I Ai
|
B
|
I BI B ou I Bi
|
AB
|
I AI B
|
O
|
ii
|
Um exemplo bem interessante e
de fácil compreensão, é a determinação da pelagem em coelhos, onde
podemos observar a manifestação genética de uma série com quatro genes
alelos: o primeiro C, expressando a cor Aguti ou Selvagem; o segundo Cch, transmitindo a cor Chinchila; o terceiro Ch, representando a cor Himalaia; e o quarto alelo Ca, responsável pela cor Albina.
Sendo a relação de dominância → C > Cch > Ch > Ca O gene C é dominante sobre todos os outros três, o Cch dominante em relação ao himalaia e ao albino, porém recessivo perante o aguti, e assim sucessivamente. |
 |
Genótipo
|
Fenótipo
|
| CC, C Cch, C Ch e C Ca |
Selvagem ou aguti
|
| CchCch, CchCh e CchCa |
Chinchila
|
| ChCh e Ch Ca |
Himalaia
|
| CaCa |
Albino
|
 |
Eventos aleatórios
Eventos como obter “cara” ao lançar uma moeda,
sortear um “ás” de ouros do baralho, ou obter “face 6” ao jogar um dado
são denominados eventos aleatórios (do latim alea, sorte) porque cada um deles tem a mesma chance de ocorrer em relação a seus respectivos eventos alternativos.
Veja a seguir as probabilidades de ocorrência
de alguns eventos aleatórios. Tente explicar por que cada um deles
ocorre com a probabilidade indicada.
|
Suponha que você jogue uma moeda
duas vezes. Qual a probabilidade de obter duas “caras”, ou seja, “cara”
no primeiro lançamento e “cara” no segundo? A chance de ocorrer “cara”
na primeira jogada é, como já vimos, igual a ½; a chance de ocorrer
“cara” na segunda jogada também é igual a1/2. Assim a probabilidade
desses dois eventos ocorrer conjuntamente é 1/2 X 1/2 = 1/4.
No lançamento simultâneo de três dados, qual a
probabilidade de sortear “face 6” em todos? A chance de ocorrer “face 6”
em cada dado é igual a 1/6. Portanto a probabilidade de ocorrer “face
6” nos três dados é 1/6 X 1/6 X 1/6 = 1/216. Isso quer dizer que a
obtenção de três “faces 6” simultâneas se repetirá, em média, 1 a cada
216 jogadas.
|
 |
Por exemplo, a probabilidade de
obter “cara” ou “coroa”, ao lançarmos uma moeda, é igual a 1, porque
representa a probabilidade de ocorrer “cara” somada à probabilidade de
ocorrer “coroa” (1/2 + 1/2 =1). Para calcular a probabilidade de obter
“face 1” ou “face 6” no lançamento de um dado, basta somar as
probabilidades de cada evento: 1/6 + 1/6 = 2/6.
Em certos casos precisamos aplicar tanto a
regra do “e” como a regra do “ou” em nossos cálculos de probabilidade.
Por exemplo, no lançamento de duas moedas, qual a probabilidade de se
obter “cara” em uma delas e “coroa” na outra? Para ocorrer “cara” na
primeira moeda E “coroa” na segunda, OU
“coroa” na primeira e “cara” na segunda. Assim nesse caso se aplica a
regra do “e” combinada a regra do “ou”. A probabilidade de ocorrer
“cara” E “coroa” (1/2 X 1/2 = 1/4) OU “coroa” e “cara” (1/2 X 1/2 = 1/4)
é igual a 1/2 (1/4 + 1/4).
|
 |
 |
Vimos que, em alguns
casos, os genes se manifestam com fenótipos bem distintos. Por exemplo,
os genes para a cor das sementes em ervilhas manifestam-se com fenótipos
bem definidos, sendo encontradas sementes amarelas ou verdes. A essa
manifestação gênica bem determinada chamamos de variação gênica descontínua, pois não há fenótipos intermediários.
Há herança de características, no entanto,
cuja manifestação do gene (também chamada de expressividade) não
determina fenótipos tão definidos, mas sim uma gradação de fenótipos. A
essa gradação da expressividade do gene, variando desde um fenótipo que
mostra leve expressão da característica até sua expressão total,
chamamos de norma de reação ou expressividade variável.
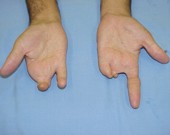
Por exemplo, os portadores dos genes para braquidactilia (dedos curto)
podem apresentar fenótipos variando de dedos levemente mais curtos até a
total falta deles.
Alguns genes sempre que estão presentes se manifestam, dizemos que são altamente penetrantes. Outros possuem uma penetrância incompleta, ou seja, apenas uma parcela dos portadores do genótipo apresenta o fenótipo correspondente.
Observe que o conceito de penetrância está
relacionado à expressividade do gene em um conjunto de indivíduos, sendo
apresentado em termos percentuais. Assim, por exemplo, podemos falar
que a penetrância para o gene para a doença de Huntington é de 100%, o
que quer dizer que 100% dos portadores desse gene apresentam (expressam)
o fenótipo correspondente.
|
|
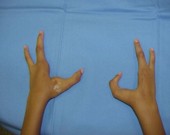
Diferentes graus de braquidactilia pela expressão variável do genótipo.
|
P
|
Planta com folhas verde claras
C c
|
|
C
Planta com folhas verde claras
c
|
CC
Verde escuro
|
Cc
Verde clara
|
Cc
Verde clara
|
cc
Inviável
|
|
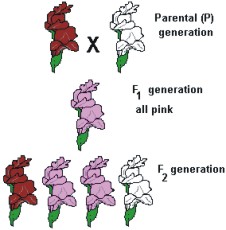
P:
|
Flor Branca
B B
|
|
|
V
Flor Vermelha
V
|
BV
cor-de-rosa
|
BV
cor-de-rosa
|
VB
cor-de-rosa
|
VB
cor-de-rosa
|
|
F1
|
Flor cor-de-rosa
V B
|
|
V
Flor cor-de-rosa
B
|
VV
Vermelha
|
BV
cor-de-rosa
|
VB
cor-de-rosa
|
BB
Branca
|
|